ミトコンドリアは運動で「数」と「質」が高まり、それが全身に適応をもたらす
ミトコンドリアは筋細胞の中でATP(エネルギー)を生み出す小さな発電所のような存在で、その量は「合成」と「分解」のバランスによって決まります。新しく作られる合成が分解を上回れば量は増え、逆に分解が優位になれば量は減少します。
運動、特に有酸素運動や高強度インターバルトレーニング(HIIT)を行うと、筋細胞内でAMPKやCaMK、p38 MAPKといったセンサー酵素が働き、PGC-1αという転写共役因子が活性化します。PGC-1αは核内の遺伝子発現を促し、ミトコンドリアDNAの複製や酸化酵素の合成を進めます。その結果、ミトコンドリアの数(量)が増えるだけでなく、酸素を利用したATP産生効率や脂質酸化能力といった質も向上します。
このとき、クエン酸合成酵素(CS)やコハク酸脱水酵素(SDH)の活性も上昇します。CSはTCA回路の入口で働き、ミトコンドリアの量や酸化代謝能力の指標となります。SDHはTCA回路の中間で機能し、電子伝達系とも直結するため、酸素利用効率や脂質酸化能力を示す重要な指標です。これらの活性化により、運動中の燃料利用は糖から脂質へとシフトしやすくなり、体内のグリコーゲンを温存する「グリコーゲン・スパリング」が起こります。その結果、長時間の運動でも疲れにくく、持久力が高まります。
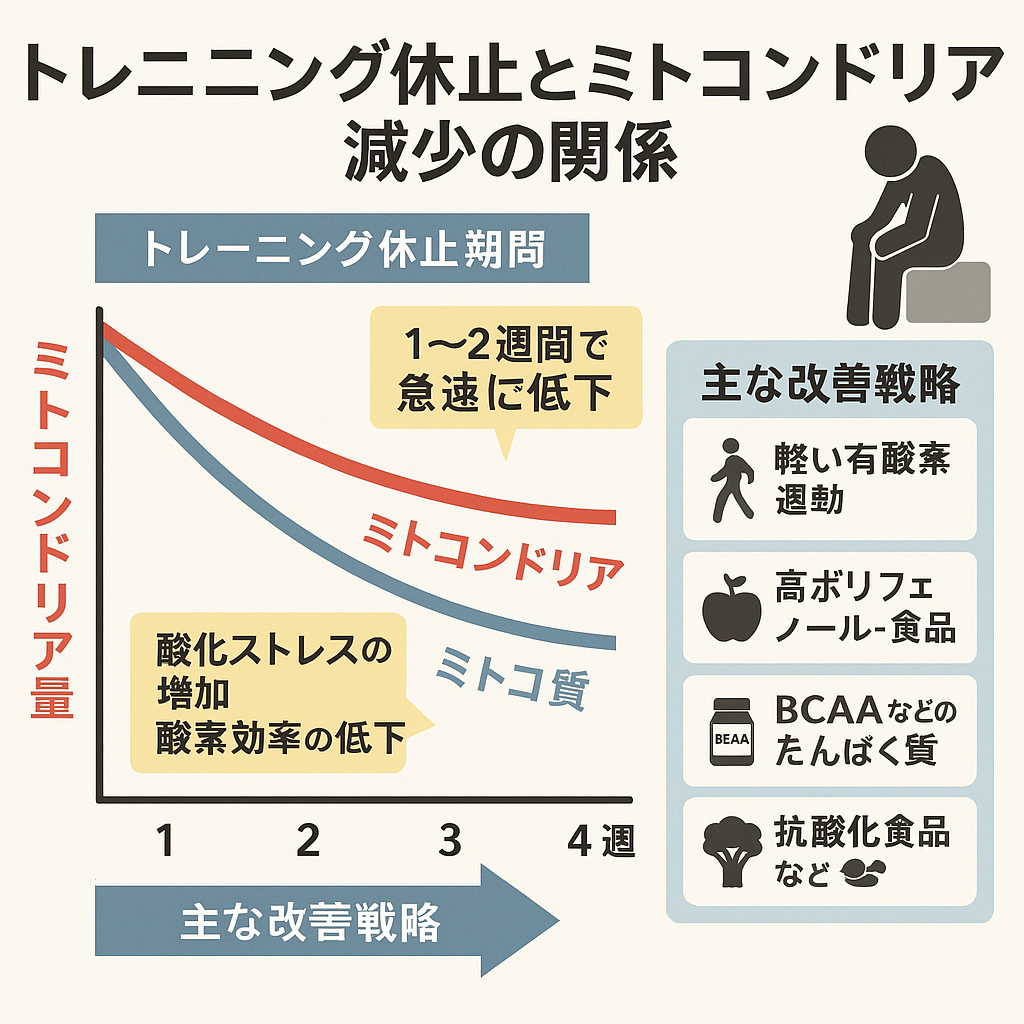
一方で運動を休止すると、PGC-1αの発現が低下し、ミトコンドリア合成が鈍ります。その間もミトファジーなどによる分解は進むため、合成より分解が優位となり、ミトコンドリアの量と質は減少していきます。CSやSDHの活性も低下し、酸素利用効率や脂質酸化能力が落ちることで糖への依存度が高まり、乳酸の蓄積が早まります。その結果、持久力や最大酸素摂取量(VO₂max)が低下し、同じ運動強度でも疲れやすくなるのです。
つまり、運動の継続は「合成>分解」の状態を維持し、ミトコンドリアの量と質を高めるために不可欠です。逆に運動をやめると「分解>合成」へと傾き、ミトコンドリア機能の低下とパフォーマンス低下を招くことになります。
運動休止とミトコンドリア機能低下のメカニズム
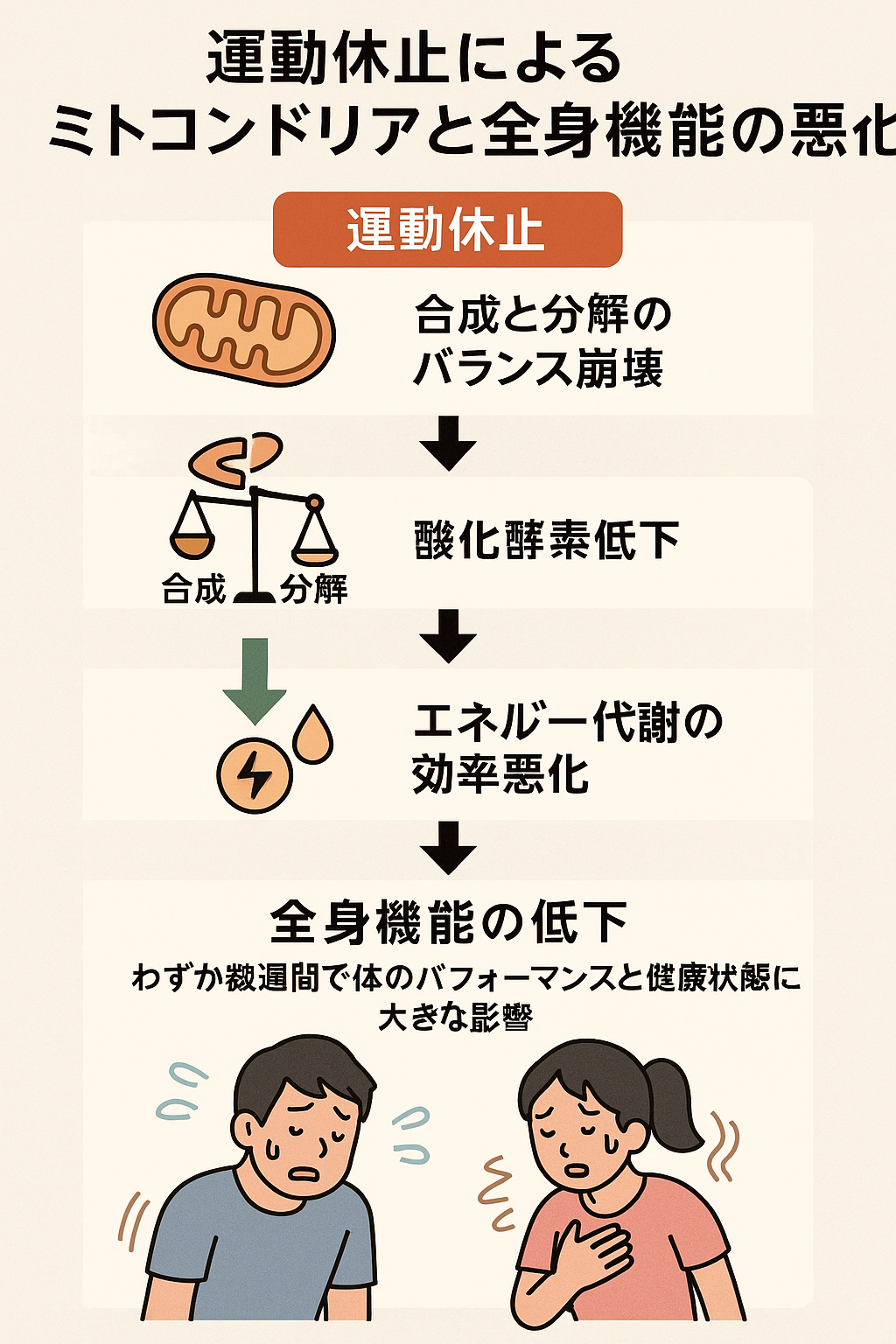
ミトコンドリアは「合成」と「分解」の均衡によって量と質が維持されています。通常、運動を続けることでAMPK・CaMK・p38 MAPKといったセンサー酵素が活性化し、PGC-1αが発現して新しいミトコンドリアが作られます。同時に、古くなったミトコンドリアはマイトファジーによって分解され、このバランスが保たれることで細胞内のエネルギー生産装置は安定しています。
しかし、トレーニングを休止するとこの仕組みが途絶えます。PGC-1αの発現が低下し合成のスイッチが切れる一方、分解は止まらず進行するため、やがて「合成<分解」となりミトコンドリアの総量は減少。これが最初の段階で起こる大きな変化です。
酸化酵素の低下と代謝効率の悪化
ミトコンドリアが減少すると同時に、エネルギー代謝を担う酸化酵素の活性も低下します。
- クエン酸合成酵素(CS):TCA回路の入口で働き、全体の酸化代謝能力の指標。
- コハク酸脱水酵素(SDH):TCA回路と電子伝達系をつなぎ、脂質酸化や酸素利用効率に直結。
- シトクロムCオキシダーゼ(COX IV):電子伝達系の最終段階で酸素を水に還元する、ATP産生の要。
これらが低下することで、脂質酸化能力が弱まり糖質依存度が上昇。結果として乳酸が早く蓄積し、同じ運動強度でも疲れやすくなります。さらに電子伝達系の効率低下によりATP産生が減り、VO₂max(最大酸素摂取量)も低下します。
実際の変化のスピード
運動をやめた場合の変化は驚くほど早く表れます。
- 休止12日後:CS 約17%減、SDH 約19%減、VO₂max 約7%低下
- 休止56日後:CS 約41%減、SDH 約38%減、VO₂max 約14%低下
このように、酸化酵素活性やミトコンドリア量の減少は酸素摂取能力の低下よりも速く、かつ大きな幅で進行します。
全身への影響
この変化は単に「酸素を取り込む力」が下がるだけではなく、「酸素を利用してATPを作る細胞内装置そのものが減る」ことが最大の問題です。
- 持久力の低下
- 疲労耐性の低下、運動後の回復の遅延
- インスリン感受性の低下、脂質異常など代謝悪化
- 筋量と代謝能力の減少によるサルコペニアのリスク増大
つまり、運動休止は酸素の運搬機能と細胞内の酸素利用能力の両方を後退させ、全身のエネルギー代謝を連鎖的に悪化させる要因となります。
| 状態 | ミトコンドリアの量・質 | 酵素(クエン酸合成酵素・コハク酸脱水酵素) | 代謝の特徴 | 結果(パフォーマンス) |
|---|---|---|---|---|
| 運動(有酸素・筋トレ・HIIT) | 合成>分解 → 量が増える・質が良くなる | 活性が高まる | 酸素をよく使える、脂質を多く燃やす、糖を節約できる | 持久力アップ、疲れにくい |
| 休止(不活動) | 合成<分解 → 量が減る・質が落ちる | 活性が低下する | 酸素利用が下がる、脂質が使えない、糖に頼る、乳酸がたまりやすい | 持久力低下、疲れやすい |
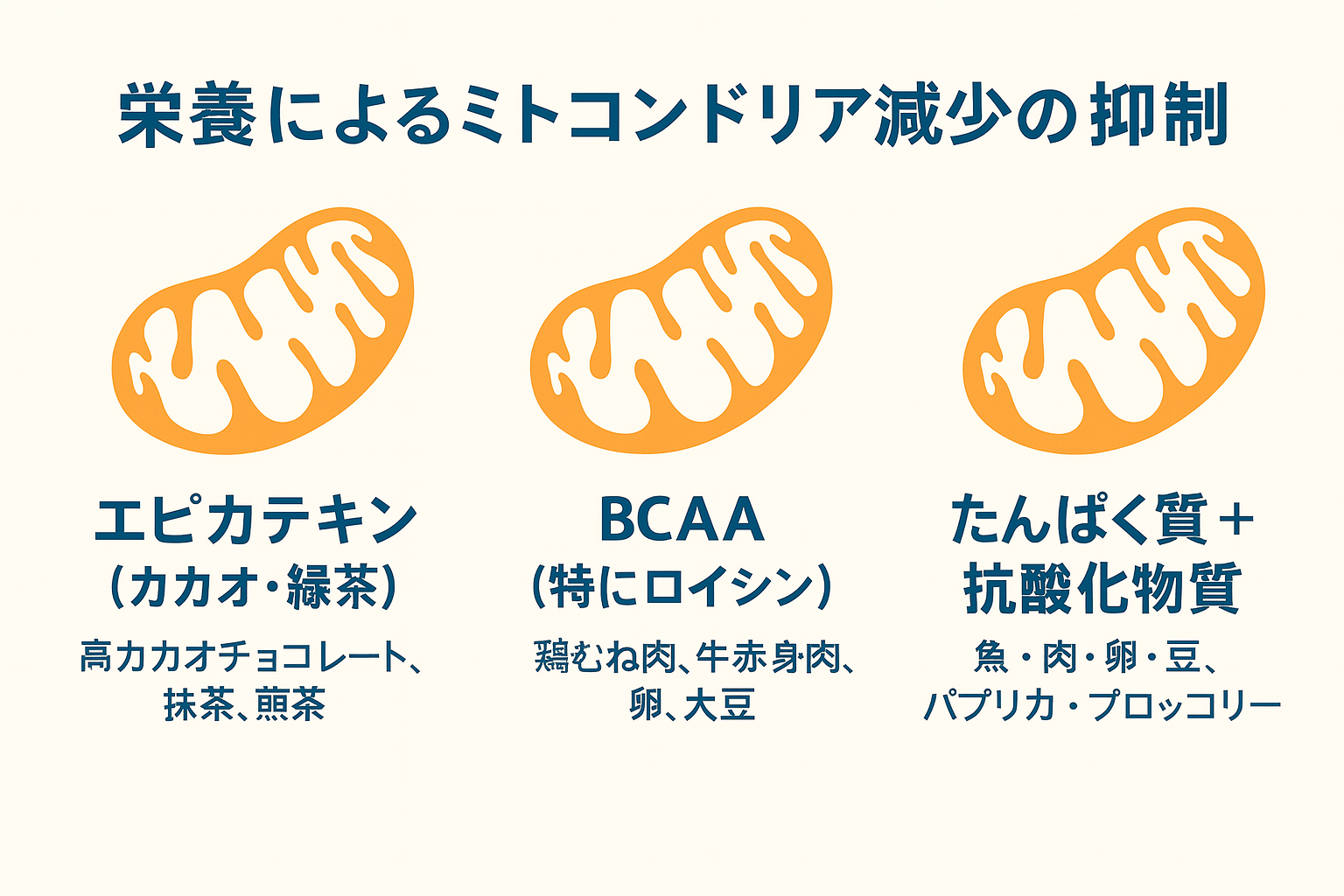
栄養によるミトコンドリア減少の抑制
運動を休止すると、まず PGC-1αの発現が低下 し、ミトコンドリアの合成が減少します。これにより、クエン酸合成酵素やコハク酸脱水素酵素、COX IVなどの酸化酵素の活性が落ち、酸素利用効率や脂質酸化能力が低下します。さらに、新生が減少した分だけ分解が優位となり、ミトコンドリア量は加速度的に減少していきます。
この悪循環を食い止めるには、以下の 3方向からの同時アプローチ が重要です。
- エピカテキン(カカオ・緑茶)
- BCAA(特にロイシン)
- たんぱく質+抗酸化物質

エピカテキン(カカオ・緑茶由来)
エピカテキンはカカオや緑茶に含まれるポリフェノールの一種で、AMPKやSIRT1を活性化し、PGC-1αの発現を促します。これにより、運動休止中でもミトコンドリア新生シグナルをある程度維持でき、CSやSDH、COX IVといった酸化酵素の発現低下を緩やかにします。結果として、酸素利用効率や脂質酸化能力の低下を遅らせることが可能になります。
主成分:フラバノール(ポリフェノールの一種)
主な作用経路:AMPK活性化、SIRT1活性化 → PGC-1α発現促進
- カカオや緑茶、リンゴなどに多く含まれるフラバノールの一種。
- 主な作用:
- PGC-1αの活性化
→ ミトコンドリア新生を促進する転写共役因子。運動と同様にミトコンドリア合成を刺激。 - eNOS(内皮型一酸化窒素合成酵素)の活性化
→ 血流改善により筋細胞への酸素・栄養供給が向上。 - 抗酸化作用
→ ミトコンドリアDNAや膜を酸化ストレスから保護。
- PGC-1αの活性化
- 人の研究では、短期間で筋力や持久力の維持・改善効果が報告されており、運動休止期にも有用と考えられます。
エピカテキン摂取例
高カカオチョコ(カカオ70%以上)、抹茶、緑茶、リンゴ皮つき。
目安:エピカテキン換算で1日50〜100mg程度。
| 段階 | 運動休止時の変化 | エピカテキンの影響 |
|---|---|---|
| PGC-1α発現低下 | 運動刺激が途絶えAMPK・CaMK経路が働かず、PGC-1α発現量が急減 | AMPK/SIRT1経路を介してPGC-1α発現を部分的に回復し、新生シグナルの低下を緩和 |
| ミトコンドリア合成量の減少 | 新生が鈍化し、合成<分解になる | PGC-1α経由でNRF-1/2、TFAMなど下流遺伝子を活性化 → mtDNA複製・酸化酵素発現増加 |
| 酸化酵素活性低下 | CS・SDH・COX IVなどの発現が減る | 酸化酵素の遺伝子発現を促進し、活性低下を遅らせる |
| 分解優位化 | 新生が減り、マイトファジーによる分解が相対的に優位に | 完全には防げないが、新生速度を高めることで分解とのバランスを改善 |
BCAA(特にロイシン)
BCAAは特にロイシンがmTOR経路を活性化し、筋タンパク質とミトコンドリア構成タンパク質の合成を促します。これにより、CSやSDHといった酵素の量的低下を抑え、酸化能力の維持に貢献します。また、mTOR活性化はオートファジーやマイトファジーの過剰活性化を抑え、ミトコンドリア分解の速度を緩めます。結果的に、筋量の維持とともにミトコンドリア総量の減少も抑制できます。
主成分:ロイシン、イソロイシン、バリン
主な作用経路:mTORC1経路活性化 → タンパク質合成促進、分解抑制
- 特にロイシンはmTOR経路を介して筋タンパク質合成を刺激。
- 主な作用:
- 筋タンパク質の分解抑制
→ 筋肉量の減少を防ぎ、ミトコンドリアを含む筋細胞の構造維持に寄与。 - エネルギー基質として利用
→ 運動時や低糖質時にエネルギー供給をサポート。 - ミトコンドリア分解(マイトファジー)の抑制
→ 筋タンパク質の劣化や過剰な分解が抑えられ、機能的なミトコンドリアが残りやすい。
- 筋タンパク質の分解抑制
- トレーニング休止期の摂取で、筋萎縮や代謝低下の進行を緩やかにできる可能性があります。
BCAA摂取例
パウダー・カプセル、または高BCAA食品(鶏むね肉、カツオ、マグロ、卵)。
目安:体重1kgあたり0.05〜0.1g(例:60kgなら3〜6g/日)。
| 段階 | 運動休止時の変化 | BCAAの影響 |
|---|---|---|
| PGC-1α発現低下 | 直接的影響は少ない | 運動時ほどではないが、筋量維持によりPGC-1α発現の低下を間接的に緩和 |
| ミトコンドリア合成量の減少 | 合成の材料(アミノ酸)が不足しやすくなる | ロイシンが筋タンパク質とミトコンドリア構成タンパク質の合成を促進し、新生をサポート |
| 酸化酵素活性低下 | CS・SDHなど構造タンパク質が減少 | 酵素タンパク質の分解抑制により活性低下を遅らせる |
| 分解優位化 | マイトファジーやユビキチン-プロテアソーム系が活性化 | mTOR活性化によりオートファジー抑制、ミトコンドリアや筋タンパク質の分解速度を低下 |
たんぱく質+抗酸化物質
良質なたんぱく質は、ミトコンドリアや酸化酵素の構造を作る材料を供給します。運動休止中も十分なたんぱく質を摂ることで、修復能力を維持できます。さらに、ビタミンCやビタミンE、ポリフェノール、カロテノイドなどの抗酸化物質は、活性酸素(ROS)によるミトコンドリアや酵素の酸化損傷を防ぎます。これにより、電子伝達系やATP合成酵素の機能低下を抑え、分解対象となる損傷ミトコンドリアの発生を減らします。
主成分:必須アミノ酸、ビタミンC・E、ポリフェノール、カロテノイドなど
主な作用経路:構造材料供給+酸化ストレス防御
- タンパク質の役割
ミトコンドリアを構成する酵素や膜タンパク質の合成材料となる。不足すると新生も修復も停滞。 - 抗酸化物質の役割
ビタミンC・E、カロテノイド(βカロテン、リコピンなど)が、ROSによる酵素・膜・DNAの損傷を抑制。これにより機能を長期維持。
| 段階 | 運動休止時の変化 | たんぱく質+抗酸化物質の影響 |
|---|---|---|
| PGC-1α発現低下 | 酸化ストレス増加がPGC-1α発現をさらに阻害 | 抗酸化作用でROSを減らし、PGC-1αの酸化的抑制を緩和 |
| ミトコンドリア合成量の減少 | 合成の材料不足、酸化損傷で合成効率低下 | 必須アミノ酸が構造材料を供給し、合成効率を維持 |
| 酸化酵素活性低下 | 酵素タンパク質が酸化変性しやすくなる | 抗酸化物質が酵素の酸化損傷を防ぎ、活性維持に貢献 |
| 分解優位化 | 損傷ミトコンドリアが増え分解が促進 | 酸化損傷を減らすことで、分解対象となるミトコンドリアを減らす |
- エピカテキン → 新しいミトコンドリアの合成促進(攻め)
- BCAA → 既存ミトコンドリアと筋細胞の分解抑制(守り)
- この組み合わせにより、運動休止期でもミトコンドリア量の維持とエネルギー代謝の安定化が期待できます。
3つの戦略を組み合わせる意義
- エピカテキンがPGC-1α経路を活性化し、新生低下を防ぐ
- BCAAが分解抑制と構造維持を行い、量の減少を防ぐ
- たんぱく質+抗酸化物質が損傷を防ぎ、機能低下を抑える
この3つを同時に行うことで、運動休止中に起こる
「PGC-1α低下 → 新生減少 → 酸化酵素低下 → 分解優位化」
という悪循環を複数の段階で食い止められます。その結果、休止期間中でもミトコンドリアの量と機能の低下を最小限に抑え、再開時に持久力や酸化的代謝能力をすばやく回復しやすくなります。
\ 最新情報をチェック /
コメント